Introducing MatPopMod, a Python package for working with matrix population models
by Owen Jones on Nov 11, 2021Guest post by François Bienvenu (Institute for Theoretical Studies, ETH Zürich)
We are happy to announce the first release of matpopmod, a Python library for the study of matrix population models.
Matpopmod provides functionalities similar to that of several R libraries (popdemo, popbio, Rage and Rcompadre): it can be used to load models from the COMPADRE / COMADRE databases; compute demographic descriptors (reproductive values, elasticities, net reproductive rate, etc); simulate deterministic and stochastic trajectories; and produce publication-ready plots.
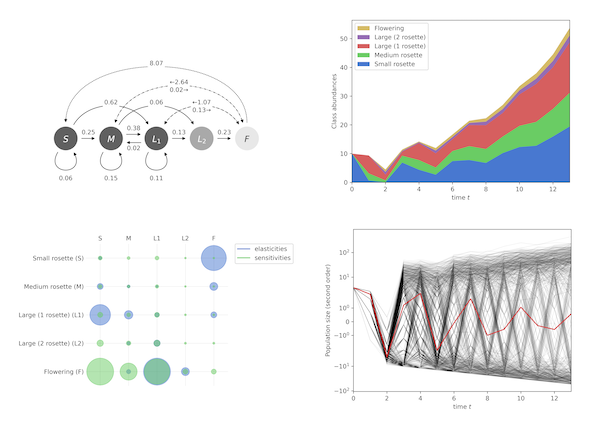
The main difference between matpopmod and existing R libraries is that matpopmod is written in Python, and primarily aimed at Python users. However, it is possible to use matpopmod with R and there are some differences in functionalities that can make matpopmod of interest even to R users. For instance, when a descriptors has several mathematical definitions (as can be the case, e.g, for the net reproductive rate or the generation time), matpopmod implements each of them, allowing you to choose the most suitable one. To find out more, you can check out the complete list of descriptors implemented in matpopmod.
Although this first release has a stable interface and already implements all of the core functionalities needed to study matrix population models, matpopmod is a work in progress and we have a lot planned for future releases. This includes fully customizable individual-based simulations; kinship estimation; quantitative assessment of the impact of within-class heterogeneity of the vital rates... and more!
If we have piqued your interest and you would like to get a more "concrete" sense of what matpopmod can do – or even give it a try! – we recommend checking our quickstart manual.
Feedback and contributions are welcome on our development repository.
Social Media
